Re salut, j'ai fini la lecture de l'article originel. Je pense que le mieux est de mettre toutes les remarques critiques dans un topic séparé, car c'est bien le genre d'articles typique de sophismes et de blablatisme inutile.
En fait, toute leur argumentation sur la position du fémur et ses incidences sur la respiration avienne versus crocodilienne, ainsi que leur remise en cause de la phylogénie theropodienne des oiseaux, ne repose que sur ce point-ci (on le trouve résumé en milieu de page), le reste n'étant que du blabla laborieux sur des considérations cardiovasuclaires crocodiles versus oiseaux, afin d'évaluer la "
respiration like" des theropodes :
 Fig. 5. Ostrich (Struthio) and Tyrannosaurus rex. Note the broadly open abdominal lumbar regions in both taxa (asterisks). In the ostrich, the semihorizontal femoral complex is incorporated into the lateral body wall and helps prevent paradoxical ventilatory collapse of the large abdominal air sacs. No such collapse-preventing mechanism existed in Tyrannosaurus: during locomotion in theropods, the distal femur rotated freely and the femoral complex was not incorporated into the abdominal wall. (Figures modified from Bolzan, 2001; Figures modified from Osborn, 1916).
Fig. 5. Ostrich (Struthio) and Tyrannosaurus rex. Note the broadly open abdominal lumbar regions in both taxa (asterisks). In the ostrich, the semihorizontal femoral complex is incorporated into the lateral body wall and helps prevent paradoxical ventilatory collapse of the large abdominal air sacs. No such collapse-preventing mechanism existed in Tyrannosaurus: during locomotion in theropods, the distal femur rotated freely and the femoral complex was not incorporated into the abdominal wall. (Figures modified from Bolzan, 2001; Figures modified from Osborn, 1916). Toute leur argumentation a comme piliers ces deux X sur les schémas, c à dire le rôle du fémur comme prévention de leur "
paradoxical ventilatory collapse of the large abdominal air sacs".
Un premier problème, c'est que leur schéma de dinosaure est dépassé depuis plus de 30 ans. Actuellement, on sait que les dinos bipèdes du genre typique allosaure, se tenaient la queue bien horizontale, le long de l'axe d'un corps et colonne vertébrale bien horizontaux.

Cette position change l'angle du fémur par rapport à l'axe de la colonne vertébrale, à la manière des oiseaux, et par conséquent rejoint les théropodes bipèdes non aviens, en similitude dynamique, aux oiseaux, y compris pour ces points-là - même si la cuisse des théropodes non aviens était plus mobile et sollicitée pour la locomotion.
Les sacs aériens des dinosaures étaient par ailleurs probablement innombrables, même localisés pneumatiquement entre les vertèbres.
(voir schémas ici :
http://www.snf.ch/SiteCollectionDocumen ... 0_21_f.pdf ).
Donc, cette comparaison anatomique-dynamique, entre un oiseau et un dinosaure à queue pendante, qui traîne au sol, est une plaisanterie qui a du plomb dans l'aile, et se réfute par une simple photo.
On peut imaginer, pourquoi pas, que la locomotion du théropode non avien participait dans une alternance droite/gauche aux mouvements côtes-sacs aériens abdominaux, accélérant leur circulation au pro rata de la cadence de la marche ou course... (idée vendue en exclusivité sur rationalisme.org ! ).
Autre chose, au sujet de cet autre comparaison "coeur des crocodiliens versus coeur des oiseaux", que les auteurs de cet article expliquent très mal, ce pour proposer celui des théropodes plutôt comme "
crocodile like" que comme "
birds like". Par une analogie foireuse de similitudes coeur crocodile - coeur des sauropsides (?).
 Fig. 1. Diagrams of the heart and great vessels in crocodilians (left) and birds (right). Both possess fully subdivided, four-chambered hearts. Crocodilians, like many sauropsids, retain paired aortae which allow mixing of oxygenated and deoxygenated blood (modified from Goodrich, 1930).
Fig. 1. Diagrams of the heart and great vessels in crocodilians (left) and birds (right). Both possess fully subdivided, four-chambered hearts. Crocodilians, like many sauropsids, retain paired aortae which allow mixing of oxygenated and deoxygenated blood (modified from Goodrich, 1930). Ce qui précède est encore un extrait du texte, sous un schéma comparatif coeur de croco - coeur d'oiseau. Leur résumé ici est plus étayé par la suite, par des éléments corrects sur le coeur de croco, mais incomplets, et aux conclusions fausses.
Voici pourquoi : Si le coeur du crocodile a bien un vaisseau - le foramen de panizza - faisant se rejoindre circulation artérielle et sanguine, ce n'est pas là le signe d'un caractère archaïque chez le crocodile, mais d'une spécialisation au mode de vie en apnée. En fait, le coeur du crocodile, à 4 cavités et 2 ventricules cloisonnés lui aussi, est tout à fait similaire et homologue à celui des oiseaux à tous points de vue,
modulo ce court foramen inter artères et une valvule (*) qu'il possède, lui permettant de bloquer complètement l'afflux de sang vers les poumons lors de la plongée (valvule) + faire dériver sa circulation sanguine (via foramen) afin de contourner les poumons. Ca lui permet de rester sous la flotte quelques heures sans respirer, d'exploiter au maximum sa réserve d'oxygène, probablement tout en conservant ses ressources musculaires. Au contraire d'autres animaux à respiration aérienne secondairement adaptés à la plongée, qui usent d'autres systèmes pour économiser énergie et oxygène lors des apnées, comme la non irrigation des organes non vitaux, le crocodile a "inventé" un coeur unique, lui permettant de changer (switcher) "sa circulation" - ce par
le coeur le plus COMPLEXE connu à l'heure actuelle, et par une circulation hybride, pouvant à la demande cloisonner circulation artérielle-sanguine, ou la mélanger.
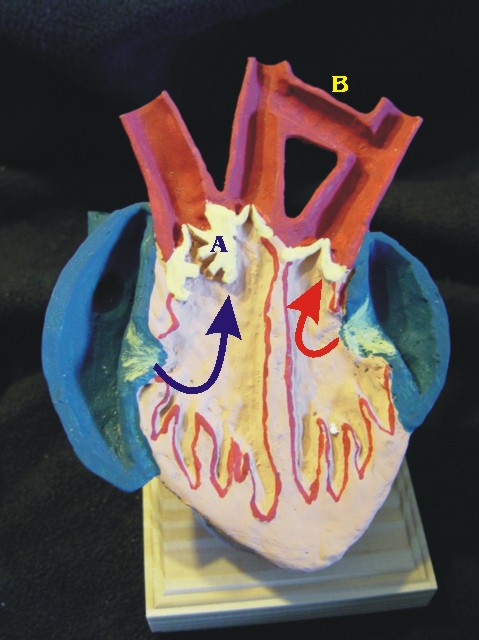
A = valve , B = formen de Panizza
(*) Les auteurs de cet article ont oublié (omis ?) de parler de cette valvule (A) qui fonctionne en coordination avec le foramen de Panizza (B), sous commande d'afflux d'adrénaline semblerait-il, et qui change la donne sur le coeur de crocodile. Circulation
hybride, selon s'il plonge ou qu'il respire, qui doit être vue comme une hyperadaptation, spécialisation donc, aux longues très longues apnées. Et non pas comme le vestige d'un caractère ancestral reptilien, car la structure du coeur de crocodile est archosaurienne, à la base donc 4 cavités et les 2 circulations cloisonnées.
Comment peut-on affirmer que cette particularité du coeur de croco n'est pas un vestige de coeur reptilien ?
C'est simple : le mélange sang artériel/veineux, constant chez les serpents/lézards, se fait à l'
intérieur du coeur, via leurs 2 ventricules g/d qui ne sont pas entièrement séparés. Or chez le crocos comme chez les oiseaux, les 2 ventricules sont entièrement cloisonnés, et seul ce foramen de Panizza, localisé
hors du coeur, faisant se rejoindre deux grosses artères, permet sa circulation si complexe. La tuyauterie du croco est basiquement celle d'un archosaurien, avec quelques ajouts.
ps :
A voir le peu de dinosaures adaptés à la vie marine ou totalement aquatique (aucun à ma connaissance) on peut en déduire que les dinosaures n'avaient pas développé ce système unique du coeur des crocos, hyper spécialisé et non pas archaïque, mais bien plutôt un coeur "
oiseaux like" ... Ce qui réfute encore toutes leurs conclusions secondaires, basées sur l'analogie dinos-crocos.
Mis à part ces points que j'ai un peu développés, l'article tente de démontrer un
impératif fémur-respiration, mais ne démontre en fait qu'une
relation mécanique, et avec bien peu d'éléments mathématiques et formules physiques (même quasiment rien), pourtant indispensables à une démonstration de fonctionnement mécanique qui se respecte.
Pour se faire une bonne idée de ce qu'est un bon article de biologie-physique-mécanique, versus celui de Ruben, consulter Google et chercher par exemple un article sur le développement des plumes de l'oiseau, voire sur la formation des plis cutanés p.e. Dans ces derniers, la physique mécanique prendra largement le pas quantitativement dans l'article, ce qui est la moindre des choses.





 Juste un aparté et rappel (pour moi-même et les 2,43° crétins qui comme nous s'intéressent aux doigts des dinosaures et suivent peut-être distraitement cette discussion ) : ce problème du frame shift, s'il est un détail simple à expliquer sur un schéma, est un des plus trompeurs qui soit - et se pige dans un autre sens d'une semaine à l'autre. C'est un peu comme une illusion d'optique, mais à recentrer en numérotant ses propres doigts avec un stylo.
Juste un aparté et rappel (pour moi-même et les 2,43° crétins qui comme nous s'intéressent aux doigts des dinosaures et suivent peut-être distraitement cette discussion ) : ce problème du frame shift, s'il est un détail simple à expliquer sur un schéma, est un des plus trompeurs qui soit - et se pige dans un autre sens d'une semaine à l'autre. C'est un peu comme une illusion d'optique, mais à recentrer en numérotant ses propres doigts avec un stylo. 













 Encore mieux car étonnant et peu connu : se munir d'une bonne loupe d'horloger et observer attentivement au niveau du talon - le lieu où commencent les plumes des pattes et où "finit" la partie à écailles (entre guillemets car ça ne finit même pas, ça donne lieu à des plumes tout simplement) - ces quelques mm de frontière cellulaire entre écailles et plumes, où on voit comment les différentes couches formant les écailles externes se transforment progressivement en plumes ; ce sur quelques mm de tissu !! La frontière entre couche/amas donnant plumes, et couche/amas formant écailles n'est pas nettement tranchée chez l'oiseau, et quelques "plumes en ébauche, encore mi-écaille" se voient dans cette zone frontalière. Bien pratique pour y observer une mosaïque de caractères sur une petite surface tégumentaire de "transition" (*).
Encore mieux car étonnant et peu connu : se munir d'une bonne loupe d'horloger et observer attentivement au niveau du talon - le lieu où commencent les plumes des pattes et où "finit" la partie à écailles (entre guillemets car ça ne finit même pas, ça donne lieu à des plumes tout simplement) - ces quelques mm de frontière cellulaire entre écailles et plumes, où on voit comment les différentes couches formant les écailles externes se transforment progressivement en plumes ; ce sur quelques mm de tissu !! La frontière entre couche/amas donnant plumes, et couche/amas formant écailles n'est pas nettement tranchée chez l'oiseau, et quelques "plumes en ébauche, encore mi-écaille" se voient dans cette zone frontalière. Bien pratique pour y observer une mosaïque de caractères sur une petite surface tégumentaire de "transition" (*). 


